Paper A4: The Ascent of Man: From Particle to Body
The material thread: from the first particles with mass through nucleosynthesis, the stars and the elements, the Earth, the origin of life and the long microbial epoch, the complex cell, the rise of animals, the move onto land, the mammalian and primate lines, to the australopithecine grade and the genus Homo — then past biology, through the peopling of the world and the genealogical line of Israel with its historicity debates named, to one ordinary human body, born in Roman Judea.
Confidence — Math: — (not engaged) — no new mathematics. Science: inference — the material thread to current science (nucleosynthesis and the elements, the origin of life and the long microbial epoch, the rise of animals and the primate line, the genus Homo, the peopling of the world), with the historicity debates of the Hebrew genealogical line named where they stand. Theology: — (not engaged) — the line is followed to one body in Roman Judea, no theological claim asserted here.
Abstract
Nothing in this paper was witnessed; every date is reconstructed — from ancient light, radioactive decay, isotopes, genomes, and bone. The previous paper closed at the electroweak vacuum, the trillionth of a second when the Higgs field switched on and matter acquired mass. This one begins there and follows the material thread forward without a break: nucleosynthesis and the light elements, the first stars and the forging of the heavier ones, the assembly of the Earth, the origin of life and the long microbial epoch, the complex cell, the rise of animals, the move onto land, and the mammalian and primate lines — to the australopithecine grade, bipedal in body and ape-grade in mind, where the animal climb reaches its last rung. The major uncertainties are flagged where they stand: the unsolved mechanism of baryogenesis, the lithium problem, the unsettled origin of life, and the molecular-versus-fossil dating gap that runs through the second half and is widest at the human–chimpanzee split. From there the body’s thread runs on through the genus Homo, the peopling of the world, and the genealogical line of Israel, to one ordinary human body born in Roman Judea.

“There is grandeur in this view of life… from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.” — Charles Darwin, On the Origin of Species (1859)
1. Introduction: reading the deep past

Nothing in this paper was witnessed. Every date and every transition in it has been reconstructed — inferred from the spectrum of ancient light, the decay of radioactive nuclei in zircon crystals, the isotopic shadow that living chemistry leaves in carbon and sulfur, the branching statistics of genomes, and the bones and footprints pressed into sediment. That this reconstruction is possible at all, and that independent methods agree across such spans of time, is one of the quieter achievements of the species. The age of the universe, 13.8 billion years (13.787 ± 0.020 Gyr; Planck Collaboration 2020), is now known to better than a percent. The age of the Earth, 4.54 billion years, is fixed by the radiometric clock in the oldest solids of the solar system to within a few hundred-thousandths.
The previous paper closed at the electroweak vacuum — the moment, about a trillionth of a second after the beginning, when the Higgs field switched on and the particles of matter acquired mass. This paper begins there and follows the material trail forward without a break: through nucleosynthesis, the first stars, the forging of the elements, the assembly of a planet, the origin of life, the long microbial epoch, the invention of the complex cell, the rise of animals, the conquest of land, the mammalian inheritance, and the primate line — ending at the last animal that stood between the apes and us. It is one continuous physical story, and the human being is its provisional latest chapter, not its purpose stated in advance.
A word on method and confidence. The deep cosmological and geological dates are firm. The biological dates carry a recurring tension: molecular clocks, which count mutations, usually place divergences earlier than the oldest known fossils, because fossils give only minimum ages and because directly measured (pedigree) mutation rates differ from long-term substitution rates. This molecular-versus-fossil gap is the single uncertainty that runs through the second half of the paper, and it is widest at the very end, at the human–chimpanzee split. It is flagged throughout rather than smoothed over.
2. The first microseconds: mass, matter, and the asymmetry that let anything remain
The story the previous paper handed forward begins with the universe already expanding and cooling out of an unimaginably hot, dense state. At about 10⁻¹² seconds, when the temperature had fallen to roughly 160 GeV (some 10¹⁵ K), the Higgs field acquired its nonzero vacuum expectation value, breaking the electroweak symmetry and giving mass to the W and Z bosons and, through their Yukawa couplings, to the quarks and leptons. It is worth stating precisely, because it is often described loosely: in the Standard Model with the measured 125 GeV Higgs, this electroweak transition is not a sharp phase transition but a smooth crossover — there is no boiling, no bubble nucleation (lattice studies; Mazumdar & White 2019). The same is true of the quark–hadron (QCD) transition near 10⁻⁶ seconds, when the quark–gluon plasma cooled below about 156 MeV and quarks became permanently confined into protons and neutrons — also a crossover for the physical quark masses (Borsányi et al. 2016).
One fact from this era is the ground of everything that follows: the universe contains matter and almost no antimatter. For every billion or so matter–antimatter pairs that annihilated, roughly one extra particle of matter remained — a baryon-to-photon ratio of about 6 × 10⁻¹⁰. That residue is everything: every star and every reader is built from the tiny surplus left after the great annihilation. The dynamical origin of this asymmetry, baryogenesis, is genuinely unsolved. It requires the three Sakharov conditions (baryon-number violation, C and CP violation, departure from equilibrium), and because both Standard-Model transitions are crossovers, Standard-Model electroweak baryogenesis cannot supply enough departure from equilibrium or CP violation. The leading candidate is leptogenesis — an asymmetry generated in the decays of heavy neutrinos and converted to baryons — attractively tied to the seesaw origin of neutrino masses (Trodden 1999; recent reviews 2024), but it remains unproven. That the universe is made of something rather than nothing is, at the level of mechanism, still an open problem.
3. The first three minutes: the light elements
Between roughly ten seconds and twenty minutes, while the universe was a cooling nuclear furnace at 10⁹ K, the free protons and neutrons fused into the lightest nuclei. With the neutron-to-proton ratio frozen near one to seven, essentially all the neutrons ended up locked in helium-4 (a mass fraction of about 0.247), with trace amounts of deuterium, helium-3, and lithium-7. Because no stable nucleus of mass 5 or 8 exists, the chain stalls at lithium; the heavier elements had to wait for stars. This Big Bang Nucleosynthesis is one of the great quantitative successes of cosmology: given the baryon density measured independently from the microwave background, the predicted abundances are fixed with no free parameters, and the deuterium prediction matches observation to about one percent (Fields et al. 2020).
There is one persistent crack. The predicted abundance of lithium-7 is about three times higher than what is measured in the atmospheres of the oldest, most metal-poor stars — a four-to-five-sigma discrepancy known as the lithium problem (Fields 2011). The community increasingly favors an astrophysical resolution: lithium being gradually destroyed by mixing inside those ancient stars, rather than any flaw in the cosmology. But the problem is not formally closed, and it is honest to name it as the one place where the otherwise excellent theory of the first three minutes still disagrees with the sky.
4. Recombination, the microwave background, and the dark ages
For its first 380,000 years the universe was an opaque plasma — photons scattering endlessly off free electrons. Then, at a temperature near 3000 K, electrons and protons combined into neutral hydrogen, the fog lifted, and the light streamed free. That light is still arriving: the cosmic microwave background, a near-perfect 2.725 K blackbody stippled with temperature fluctuations of about one part in 100,000 (Hu & Dodelson 2002). Those fluctuations are the seeds of all later structure — the faint overdensities that gravity would spend the next hundreds of millions of years pulling into the first stars and galaxies. The CMB is the most precisely measured object in cosmology, and its pattern of acoustic peaks fixes the geometry of the universe as flat and pins down its contents.
After recombination came the cosmic dark ages: a long, cold, structureless interval of neutral hydrogen, lit by nothing but the redshifting afterglow, lasting until gravity finally assembled objects dense enough to ignite. (The chief unresolved tension in modern cosmology, the roughly five-sigma disagreement between the early-universe and locally measured values of the expansion rate — the Hubble tension, recently sharpened rather than dissolved by JWST; Riess et al. 2024 — sits adjacent to this story but does not change its sequence.)
5. The first light: stars, galaxies, and reionization

Within the densest knots of dark matter — halos of perhaps a hundred thousand to a million solar masses — primordial gas, cooled inefficiently by molecular hydrogen, collapsed and ignited the first stars, the metal-free Population III, somewhere around 100 to 250 million years after the beginning. Lacking the cooling that heavier elements provide, these first stars are thought to have been massive and short-lived, tens to hundreds of times the mass of the Sun (Bromm 2013). Their ultraviolet light and their explosive deaths produced the first heavy elements and began to reionize the surrounding hydrogen; as galaxies assembled and grew, their radiation carved expanding bubbles of ionized gas that merged and, by about a billion years after the beginning, had reionized the universe entire (Robertson 2022).
This is the most rapidly moving frontier in the whole sequence. The James Webb Space Telescope has now spectroscopically confirmed galaxies at redshifts beyond 14 — within about 280 million years of the Big Bang (Carniani et al. 2024). And it has produced a genuine surprise: early galaxies appear brighter, more massive, and more numerous than pre-launch models of galaxy formation predicted. Whether this signals merely that early star formation was unusually efficient or top-heavy — the mainstream reading — or something deeper about the cosmological model is, as of this writing, actively debated. Population III stars themselves remain undetected; their direct signature is still being hunted.
6. Stardust: the origin of the elements
The reader is made of elements that did not exist in the early universe. Carbon, nitrogen, oxygen, phosphorus, iron — none emerged from the first three minutes, which produced only hydrogen, helium, and a trace of lithium. Everything heavier was forged inside stars and dispersed by their deaths, over the nine billion years of cosmic chemistry that preceded the Sun.
Stars fuse hydrogen to helium through the proton–proton chain and the catalytic CNO cycle; helium fuses to carbon through the triple-alpha process, and onward to oxygen, neon, silicon, and up to the iron group, where fusion can extract no more energy because iron has the highest binding energy per nucleon. Elements heavier than iron cannot be built by fusion at all; they are assembled by neutron capture — the slow s-process in aging stars, and the rapid r-process in violently neutron-rich environments, which builds the heaviest nuclei, gold and platinum and uranium and the lanthanides (Burbidge, Burbidge, Fowler & Hoyle 1957; Kobayashi, Karakas & Lugaro 2020). Core-collapse and thermonuclear supernovae scatter this enrichment into the interstellar medium, where it seeds the next generation of stars and planets. In 2017 the neutron-star merger GW170817 was caught in gravitational waves and in the light of its kilonova, giving the first direct evidence that such mergers manufacture r-process elements (Kasen et al. 2017) — though whether mergers or rare specialized supernovae dominate the heaviest-element budget remains contested (Cowan et al. 2021). The phrase is literal, not poetic: the calcium in bone and the iron in blood were made in stars that died before the Sun was born.
7. A star and its third planet
About 4.567 billion years ago — a date fixed with extraordinary precision by lead–lead dating of calcium–aluminum-rich inclusions, the oldest solids in any meteorite (Connelly et al. 2012) — a fragment of a molecular cloud collapsed into the proto-Sun, wrapped in a protoplanetary disk of gas and dust. Isotopic evidence indicates the disk’s first solids formed in under 200,000 years. Within that disk, dust grains coagulated into planetesimals, planetesimals into embryos, and embryos, through a few tens of millions of years of collisions, into planets. The Earth’s main accretion was largely complete by about 4.54 billion years ago.
Then the young Earth was struck. The giant-impact hypothesis holds that a Mars-sized body, Theia, collided with the proto-Earth and ejected the debris that coalesced into the Moon — the standard explanation for the Moon’s low density, its depletion in volatiles, and its near-identical isotopic composition to Earth’s (Canup & Asphaug 2001). The date of this impact is debated, with recent work favoring a relatively late event around 4.35 billion years ago. The energy left the Earth sheathed in a magma ocean; as it cooled, outgassing and impact delivery built a steam atmosphere that condensed into the first oceans. Remarkably, oxygen isotopes in the Jack Hills zircons of Western Australia — the oldest surviving terrestrial material, up to 4.40 billion years old (Valley et al. 2014) — indicate liquid-water oceans and habitable surface conditions within about 250 million years of the planet’s formation. The old picture of a hellish, sterile Hadean has given way to one of an early, wet, and possibly habitable world. The once-standard “Late Heavy Bombardment,” a supposed cataclysmic spike of impacts around 3.9 billion years ago, is now disfavored; the impact record looks instead like the smoothly declining tail of accretion (lunar-impactite isotope analyses, 2021).
By the close of this chapter the stage is set: a rocky planet, liquid water, an atmosphere, a steady energy supply from the Sun and from the Earth’s own hot interior, and a full inventory of the elements that chemistry can use. What happens next is the part we understand least and care about most.
“The nitrogen in our DNA, the calcium in our teeth, the iron in our blood… were made in the interiors of collapsing stars. We are made of starstuff.” — Carl Sagan, Cosmos (1980)
8. The origin of life
Somewhere between the condensation of the oceans (~4.4 Gya) and the most recent common ancestor of all living things (~4.2 Gya, below), life began. The window is narrow, which by itself suggests that the origin of life, given an Earth-like planet, was not a freakishly improbable event — though that inference rests on a sample of one.
The earliest direct evidence is contested. Robust biosignatures — stromatolites and microbial structures in the ~3.48-billion-year-old rocks of the Pilbara in Western Australia — are broadly accepted. Older claims are not: the ~3.7-billion-year-old “stromatolites” of Isua, Greenland (Nutman et al. 2016) were challenged as deformation features (Allwood et al. 2018), and the ~4.1-billion-year-old carbon in a single Jack Hills zircon (Bell et al. 2015) cannot be cleanly distinguished from non-biological carbon. The field now treats carbon-isotope signatures alone as non-diagnostic, because abiotic processes such as serpentinization can mimic the isotopic “lightness” of biological carbon (Javaux 2019). Consilience across morphology, isotopes, and sedimentary context is required, and only by ~3.5 Gya is it firmly in hand.
How life began is the largest open problem in this paper. The historically polarized debate between “replication-first” and “metabolism-first” is dissolving into an integrated systems-chemistry view. The RNA world remains the leading framework for the genetic stage: RNA can both store information and catalyze reactions (the ribosome is itself a ribozyme), so a stage in which RNA did both is attractive — but the spontaneous, non-enzymatic emergence of a self-replicating RNA from prebiotic feedstocks has not been demonstrated. Cyanosulfidic chemistry (Patel et al. 2015) shows that a single reaction network driven by ultraviolet light, starting from hydrogen cyanide and hydrogen sulfide, can yield precursors of nucleotides, amino acids, and lipids together — the strongest demonstrated route to the building blocks, and one that favors a sunlit surface setting. The alkaline hydrothermal vent hypothesis (Martin, Russell, Lane) proposes instead that life began at deep-sea vents, where natural proton gradients across thin inorganic membranes mimic the chemiosmotic energy economy that all cells still use, with carbon fixed through a proto-Wood–Ljungdahl pathway (Lane & Martin 2012; Preiner et al. 2020). And Miller–Urey chemistry, reanalyzed with modern instruments, yields far more amino acids than first reported. What is solved: plausible abiotic routes to most of the building blocks. What is not solved: the transition from prebiotic chemistry to a compartmentalized, self-replicating, evolving system — the Darwinian threshold — and which environment hosted it.
Whatever the route, it converged on a single lineage. The Last Universal Common Ancestor (LUCA) — the node from which Bacteria and Archaea diverge — has been reconstructed by molecular clocks to about 4.2 billion years ago (4.09–4.33 Gya; Moody et al. 2024), older than the oldest accepted fossils. LUCA was not the first life and not a simple proto-cell: the reconstruction gives it a genome of roughly 2.5 million base pairs and on the order of 2,600 proteins, an anaerobic metabolism fixing CO₂ with hydrogen, and — strikingly — a rudimentary antiviral immune system, implying it already lived under viral predation, inside a functioning ecosystem. By the time of the common ancestor, in other words, life was already complex, already old, and already not alone.
9. The microbial Earth: the long reign and the poisoning of the air
For most of its history, life on Earth was exclusively microbial. From LUCA, the two great prokaryotic domains — Bacteria and Archaea — diverged and spread, living by chemistry: drawing energy from hydrogen, sulfur, iron, and carbon dioxide at vents and in sediments. The earliest metabolisms were anaerobic; the atmosphere held essentially no free oxygen.
The single most consequential biological innovation in the planet’s history was oxygenic photosynthesis — the trick, evolved in the ancestors of cyanobacteria, of splitting water with sunlight and releasing oxygen as waste. Anoxygenic photosynthesis came first, using donors such as hydrogen sulfide and dissolved iron; the combination of two photosystems into a single water-splitting cell was the breakthrough (Fischer, Hemp & Johnson 2016). Its consequences were planetary. Beginning around 2.4 billion years ago, oxygen accumulated irreversibly in the atmosphere — the Great Oxidation Event (Lyons, Reinhard & Planavsky 2014) — recorded by the disappearance of a sulfur-isotope anomaly and the rusting of the oceans into banded iron formations. To the anaerobic biosphere that produced it, oxygen was a poison; the GOE was very likely the first mass extinction, an “oxygen catastrophe.” The collapse of an earlier methane greenhouse plausibly triggered the first global glaciations, the Huronian “snowball” episodes. A live debate concerns the lag: trace-oxygen “whiffs” appear hundreds of millions of years before 2.4 Gya, suggesting that oxygenic photosynthesis evolved well before the air actually changed, and that something suppressed oxygen’s accumulation for a very long time (Poulton et al. 2021). The waste product of one kind of cell rewrote the chemistry of the whole planet and set the stage for everything that breathes.
10. The complex cell
Every organism larger than a film of slime — every animal, plant, fungus, and seaweed — is built from eukaryotic cells: cells with a nucleus, an internal skeleton, membrane-bound compartments, and, crucially, mitochondria, the power plants that make large, energy-hungry cells possible. The eukaryotic cell did not arise gradually from a bacterium. It arose from a merger. An archaeal host cell — specifically a member of the recently discovered Asgard archaea, the closest known archaeal relatives of eukaryotes — engulfed or entangled an alphaproteobacterium, and the two became one: the bacterium became the mitochondrion (Eme et al. 2017). This single endosymbiosis is the founding event of all complex life. (Chloroplasts came later, in a second, independent endosymbiosis of a cyanobacterium, founding the algae and plants.)
The Asgard archaea are the key recent evidence. The first one was cultivated only in 2020 (Imachi et al. 2020), and it proved to have long cellular protrusions and, as later work showed, a genuine actin cytoskeleton (Rodrigues-Oliveira et al. 2023) — the kind of cellular machinery once thought uniquely eukaryotic. The timing is being revised older: where classic estimates placed the last eukaryotic common ancestor around 1.6–2.1 billion years ago, a 2025 molecular-clock analysis pushes eukaryotic origins back toward 3.0–2.25 billion years ago, with mitochondrial endosymbiosis already underway by ~2.2 Gya (Vosseberg et al. 2025) — a date well ahead of the unambiguous fossil record, and contested for exactly that reason. However the dating settles, the architecture is agreed: complex life is a chimera, an archaeon and a bacterium that stopped being two things.
11. Toward animals: many cells, a frozen planet, and the first large bodies
Multicellularity is not a single invention; it evolved independently more than twenty times across the tree of life — in animals, fungi, plants, red and brown algae, and others — each lineage crossing the threshold from its own unicellular ancestors. The long middle stretch of Earth history, roughly 1.8 to 0.8 billion years ago, was once called the “Boring Billion” for its apparent evolutionary and environmental stasis; recent biomarker work (Brocks et al. 2023) reveals it was in fact populated by a hidden world of early eukaryotes, invisible to standard methods — less boring than under-sampled.
It ended in ice. The Cryogenian glaciations — the Sturtian (~717–660 Mya) and Marinoan (~650–635 Mya) “Snowball Earth” episodes — buried much or all of the planet under ice. There is a growing case that this extreme environmental filter helped drive the repeated evolution of complex multicellularity. In the warm aftermath came the Ediacaran Period (635–538.8 Mya) and the first large, complex, soft-bodied organisms — the Ediacaran biota, strange fronds and quilted discs such as Dickinsonia, some of which biomarker evidence has now confirmed as genuine animals (Bobrovskiy et al. 2018). For the first time, life was visible to the naked eye, and the world was no longer microbial.
12. The Cambrian and the vertebrate body plan
Then, beginning at 538.8 million years ago (the revised Ediacaran–Cambrian boundary; ICS 2022), came the Cambrian explosion — the geologically rapid appearance, over a few million years, of nearly all the major animal body plans, the phyla, that exist today. It is not a single instant but an interval: a first phase of small shelly fossils and burrowing, then a second phase of large articulated animals, preserved spectacularly in the Chengjiang biota of China (~518 Mya) and the Burgess Shale of Canada (~508 Mya). No single cause is accepted. A modest rise in shallow-water oxygen was probably permissive rather than decisive (Stockey et al. 2024); ecological feedbacks — the onset of active predation, the churning of the seafloor by burrowers, an escalating arms race between predators and prey — are increasingly favored as the proximate engine, with the genetic toolkit for building bodies already in place. Whether the “explosion” reflects genuine rapid innovation or the sudden fossilization of lineages that had quietly diverged earlier remains debated.
Among the Cambrian animals were the first chordates and the first vertebrates — small, jawless, soft-bodied swimmers such as Haikouichthys from Chengjiang. Jawless fishes diversified through the Ordovician and Silurian. Then came jaws, evolved from the skeletal arches that had supported the front gills — one of the most consequential innovations in the animal line, opening the way to active predation at large size. A remarkable trove of early Silurian fossils from China, described in 2022, pushed the oldest complete jawed fishes back to about 436 million years ago and the oldest vertebrate teeth to ~439 Mya (Zhu et al. 2022), substantially closing the long gap between the molecular estimate for jawed vertebrates and their fossil record.
13. Onto the land

The vertebrate move from water to land, through the Devonian, is among the best-documented major transitions in the fossil record. Within the sarcopterygians — the lobe-finned fishes — the fleshy pectoral fin elaborated an internal skeleton with the precursors of the upper arm, forearm, and wrist, and weight-bearing joints. The transitional forms are famous: Tiktaalik roseae (~375 Mya), whose pectoral fin could bear weight and flex at a proto-wrist (Stewart et al. 2020), and the early digited tetrapods Acanthostega and Ichthyostega (~365 Mya), which still lived largely in water. Digits and limbs evolved before full terrestriality, for moving through shallow, weedy, oxygen-poor water rather than for walking on dry land.
The conquest of land was completed by the amniotes, whose self-contained egg — with its protective membranes and water-conserving physiology — freed reproduction from standing water and opened the dry continental interior. The oldest amniote body fossils are from the Late Carboniferous (~318 Mya), and very shortly after their origin the amniote line split into the two great branches that still divide the land vertebrates: the sauropsids, leading to reptiles and birds, and the synapsids, the branch that leads to mammals — and to us. (A 2025 report of clawed trackways in Australia dated to ~355 Mya may push this split tens of millions of years deeper into the Carboniferous than the body fossils show; if it holds, the early record is even more incomplete than assumed.) From this point, the relevant thread is the synapsid one.
14. The mammalian inheritance

The synapsids dominated the land before the dinosaurs. Through the Permian they radiated into the therapsids and then the cynodonts, in which a long series of changes — documented bone by bone — gradually assembled the mammalian body. The most celebrated is the jaw-to-ear transition: bones that had formed the reptilian jaw joint shrank, migrated, and became the tiny malleus and incus of the mammalian middle ear, while a new jaw joint formed from the dentary. Endothermy, fur, differentiated teeth, a secondary palate, and prolonged parental care were acquired piecemeal across the cynodont–mammaliaform grade (the timing of warm-bloodedness is itself debated — inner-ear evidence suggests a Late Triassic origin around 233 Mya; Araújo et al. 2022 — against tooth-growth evidence for slower, more reptile-like metabolism persisting into the Early Jurassic; Newham et al. 2020). The first true mammals appear around the Triassic–Jurassic boundary, ~205–200 Mya.
But this was not their world yet. The end-Permian extinction (~252 Mya, “the Great Dying,” driven by the Siberian Traps volcanism) had nearly ended complex life altogether — up to ~96% of marine species lost (Black et al. 2024) — and the end-Triassic extinction that followed cleared the way for the dinosaurs, who dominated the land for the next 135 million years. Through all of it, mammals stayed small, and largely nocturnal — a “nocturnal bottleneck” written into the genetics of mammalian vision. They were patient.
Their inheritance came by catastrophe. 66 million years ago, an asteroid roughly ten kilometers across struck what is now the Yucatán at Chicxulub, in the boreal spring (During et al. 2022). The impact threw enough sulfur aerosol, soot, and fine silicate dust into the stratosphere to shut down photosynthesis for around two years and darken the sky for up to fifteen (Senel et al. 2023). About three-quarters of species died, including every non-avian dinosaur. (The contemporaneous Deccan Traps volcanism was a contributing stress, but modeling favors the impact as the decisive trigger; Chiarenza et al. 2020.) Into the empty world, in the Paleocene, the surviving mammals radiated explosively — growing larger, diversifying into the modern orders, taking the daylight ecological roles the dinosaurs had vacated. The age of mammals had begun, and with it the lineage that would lead to the primates.
15. The primate line
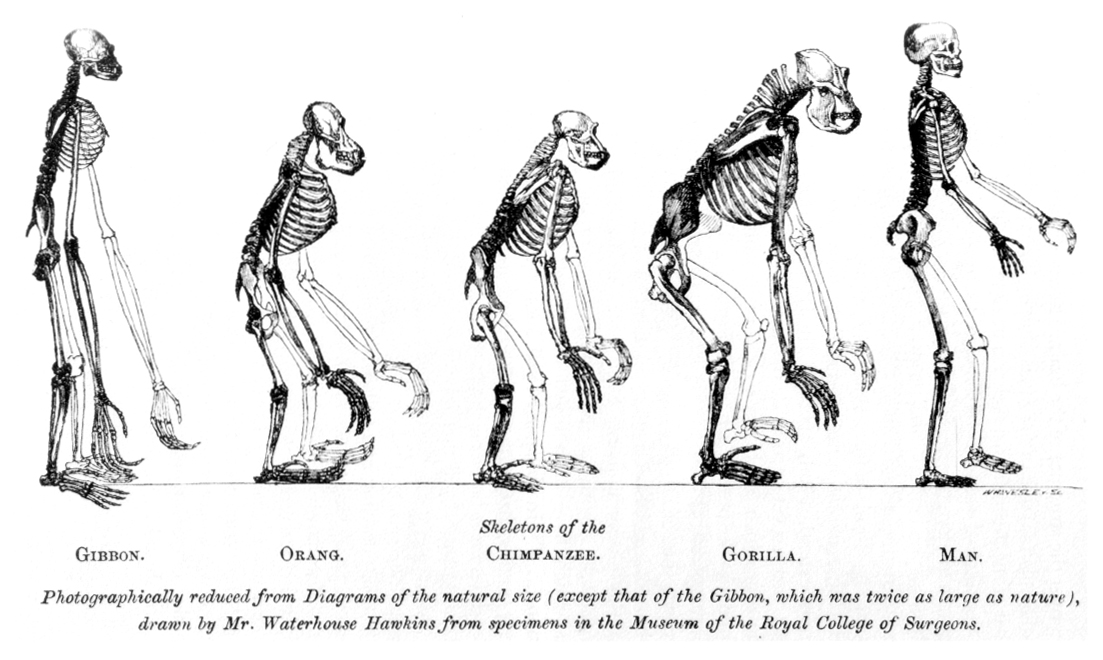
The primates emerged at or just after the K–Pg boundary. The oldest stem primates, the plesiadapiforms — small, arboreal, somewhat squirrel-like — appear within ~140,000 years of the extinction (Purgatorius; Wilson Mantilla et al. 2021), implying the lineage probably originated in the Late Cretaceous, before the impact. The first true primates split into the two living branches: the strepsirrhines (lemurs, lorises, galagos) and the haplorhines (tarsiers and the anthropoids — monkeys and apes). The defining primate features — grasping hands with nails rather than claws, forward-facing eyes with excellent depth perception, and enlarged brains — are the adaptations of an animal living by precision in the three-dimensional world of the trees.
The anthropoids arose in the late Eocene of Afro-Arabia; one branch rafted across the then-narrower Atlantic to become the New World monkeys, while the catarrhines of the Old World split, around 25–29 million years ago, into the Old World monkeys and the apes (Hominoidea). The apes radiated through the Miocene into a diversity far greater than today’s — the true “planet of the apes,” with many genera across Africa and Eurasia (Begun and others). From this radiation, the living apes branched off the line leading to humans in a settled order: the gibbons first (~17 Mya), then the orangutans (~12–16 Mya), then the gorillas (~8–10 Mya). The branching order is not in doubt, though the placement of individual Miocene fossil apes is a “neverending controversy” (Alba et al. 2023), and a 2023 find from Türkiye, Anadoluvius, has revived debate over whether the African ape and human group had Mediterranean roots (Sevim-Erol et al. 2023). One branch remained.
16. The last animal: our Uncle Monkey
The last lineage to split from ours is the one leading to the chimpanzees and bonobos. The last common ancestor of humans and chimpanzees — our Uncle Monkey, the most recent animal from which both we and our nearest living relatives descend — lived somewhere in the range of 6 to 9 million years ago. That this is a range, and a contested one, must be stated plainly. Molecular clocks based on long-term substitution rates favor the younger end (~5–7 Mya); clocks calibrated to directly measured, slower, generation-by-generation mutation rates push the split older, toward 7–13 Mya (Langergraber et al. 2012; Moorjani et al. 2016). The split was probably not a clean event but an extended separation, perhaps with gene flow continuing for a time after it began.
One point deserves emphasis, because it is the most common misunderstanding of human origins: the last common ancestor was not a chimpanzee. Chimpanzees have had exactly as long as we have to evolve, and they have their own derived specializations — knuckle-walking, thin tooth enamel, particular cranial and dental forms. They are our cousins, not our ancestors, and they are not “primitive.” What the Uncle Monkey actually looked like is genuinely uncertain. The 4.4-million-year-old Ardipithecus ramidus — “Ardi” — reshaped the question: it shows none of the knuckle-walking specializations of living African apes, suggesting the ancestor may have been a more generalized arboreal climber rather than a chimp-like form (White et al. 2009), though a competing reading of its hand argues for shared suspensory adaptations (Prang et al. 2021). The field is genuinely split.
On our side of that split stand the earliest hominins — the taxa more closely related to us than to chimpanzees. Sahelanthropus tchadensis (~7 Mya, Chad), Orrorin tugenensis (~6 Mya, Kenya), and Ardipithecus (~5.8–4.4 Mya, Ethiopia) are the candidates, and what unites them, contentiously, is the earliest evidence of bipedalism — habitual upright walking, the first and most fundamental of the changes that distinguish our line. The evidence is fragmentary and disputed; whether Sahelanthropus in particular was a habitual biped, or even a hominin at all, is argued in the literature to the present day (Daver et al. 2022, contested by Cazenave et al. 2024). Upright walking, not a large brain, came first.
By 3.9 to 2.9 million years ago, the case is unambiguous. Australopithecus afarensis — the species of the famous “Lucy” — was a committed terrestrial biped, as the 3.66-million-year-old Laetoli footprints record directly: a trail of upright walkers pressed into volcanic ash. And yet the australopithecine brain was still ape-grade, around 400–550 cubic centimeters, organized along ape-like lines, though already showing the prolonged, human-like growth that signals an extended childhood (Gunz et al. 2020). These were animals: bipedal, small-brained, living in a bushy radiation of related forms (the 2025 Ledi-Geraru fossils show up to four hominin lineages coexisting in East Africa around 2.6 Mya; Villmoare et al. 2025). They made, at most, the simplest flaked stone tools — and even toolmaking may predate our genus, given the 3.3-million-year-old flakes of Lomekwi (Harmand et al. 2015).
Here the animal climb reaches its last rung. The australopithecine grade — a creature that walked as we walk but did not yet think as we think, bipedal in body and ape-grade in mind — is the last animal state before the threshold. What crosses that threshold around 2.8 million years ago, in the lineage that becomes the genus Homo — the enlarging brain, the systematic toolmaking, and eventually the reflective, self-aware mind that could one day reconstruct the very sequence written in this paper — is the thread of the companion paper. The body’s thread, the one this paper has been following, runs on a little further.
17. The body becomes human

The body-line ran on from the australopithecines through the genus Homo, the brain enlarging, the frame settling into its modern proportions, until — by about 300,000 years ago, the date of the earliest Homo sapiens fossils from Jebel Irhoud in Morocco (Hublin et al. 2017) — there existed a body anatomically indistinguishable from the reader’s.
The material record can trace one further thing about this body, though only in its shadow: the behavior that betrays a mind. Through the African Middle Stone Age the human line began to do things that seem to need an interior to explain — grinding and engraving pigment and piercing shell beads at Blombos Cave around seventy-five to a hundred thousand years ago (Henshilwood et al. 2002); and, in the later flowering the European caves made famous, painting walls and carving figurines (Chauvet, the Hohle Fels figure, the Sulawesi panels, forty-odd thousand years old and now matched by finds across Africa and Asia), and burying the dead with evident care. Whether this marks a sudden “revolution” or — as the evidence now leans — a long, gradual, mosaic accumulation reaching back toward the body’s own origin is genuinely debated (McBrearty & Brooks 2000). It is as much of a historical account of the arrival of consciousness as the rock affords, and it comes with two honest limits: the record dates behavior, not the interior — a bead or a burial shows that a mind was at work, not what it was like to be that mind — and there is no first moment in it, no stratum where the lights demonstrably come on. What the interior is, and whether it had a first waking the rock cannot see, is the companion paper’s question.
On the material thread there is only the body — some thirty-seven trillion cells, the iron in its blood and the calcium in its bones forged in stars that died before the Sun, an organism continuous in every tissue with the long climb behind it, and continuous with every other animal in the one respect that will matter at the end: it can die. But the end is not yet. First the body is a link in a line — it would beget others, and they others — and it is the line, now, that the thread follows.
18. Out of the climb and into history
The species the last section left at three hundred thousand years did what living things do: it bred, and it spread. Sometime after about seventy thousand years ago a population carrying fully modern behavior moved out of Africa and, over tens of millennia, peopled the rest of the planet — reaching Australia by roughly fifty thousand years ago and the Americas by at least sixteen thousand, interbreeding along the way with the Neanderthals and Denisovans it met, whose DNA the reader still carries in traces. Then, beginning around twelve thousand years ago and independently in several places — the Fertile Crescent first among them — came the largest change in the human way of life since the body itself: agriculture. Settled villages replaced the moving band; stored surplus made possible specialists, then towns, then, by about five thousand years ago in Mesopotamia and Egypt, cities and writing. With writing the long silent climb acquires, for the first time, a documentary record of itself. Recorded history begins.
The thread the rest of this paper follows narrows here, and the narrowing should be named for what it is. The story so far has been the story of everything alive, then of one species. But it is going toward one body — a single human being, born at one place and one time — and to reach that body it must leave the general history of the species and follow a single line of descent: parents to children, down one chain among the billions, across the centuries from the first cities to the first century of the common era. Of all those chains, this paper follows the one that the tradition from which the body came marks as the line set apart to carry it. That line is the people of Israel.
19. The line: the people of Israel
The tradition’s account of the line begins with a herdsman called Abraham, who in the early second millennium BCE — by the tradition’s own reckoning; the patriarchal narratives were written down a great deal later, and how much history stands behind them is genuinely debated — left Mesopotamia for the land of Canaan under a promise. The promise is the reason the line is followed at all: in thee shall all families of the earth be blessed (Genesis 12:3) — a covenant that the blessing of all peoples would descend through his offspring. Through Isaac, through Jacob (renamed Israel), and through Jacob’s twelve sons the promise is carried; and within it a narrower line is marked early, the line of the fourth son, Judah, to whom the scepter is promised (Genesis 49:10). From the very start the tradition is tracing not merely a people but a line of descent inside it.
The narrative goes down into Egypt in famine, multiplies there into a people, and is delivered out of bondage in the Exodus under Moses — the constitutive memory of the nation and the setting of the Law given at Sinai. As history this is the hardest stretch to anchor: an event of the magnitude the text describes has left no direct archaeological trace, and its historicity and scale are debated; what is not in doubt is that the memory of it became the spine of a people’s identity. By about 1200 BCE a people called Israel is on the ground in the central highlands of Canaan — named in stone for the first time at about that date, the earliest the name appears outside the Bible. The inscriptions and manuscripts that attest the line from here on are gathered, as the physical record of the kept people and its text, in Paper A5.
Out of a loose confederation came a monarchy: Saul, then David around 1000 BCE, then Solomon and the first Temple in Jerusalem. The scale and splendor of that united kingdom are contested between archaeologists who read the record maximally and those who read it minimally — but that a Davidic dynasty was real is no longer in serious doubt — settled by a ninth-century inscription naming the “House of David,” read with the rest of the physical record in Paper A5. David is the figure the line bends toward, because the promise narrows onto him — an everlasting throne (2 Samuel 7) — and from here the awaited deliverer is expected to come of David’s line. The kingdom then split, Israel in the north and Judah in the south; Assyria destroyed and deported the north in 722 BCE; Judah, carrying the Davidic line, lasted another century and more.
Then Judah fell too. Babylon took Jerusalem and burned the Temple in 586 BCE and carried the people into exile — the catastrophe that, paradoxically, forged the tradition into its lasting form, as the exiles gathered and edited the scriptures that held the line’s memory. When Persia broke Babylon, Cyrus let the exiles return (from 538 BCE); a second Temple rose, the Torah was consolidated, and the people re-rooted under a succession of empires — Persian, then Greek after Alexander (332 BCE), then, after the Maccabean revolt won a brief independence (from 167 BCE) under the Hasmoneans, Roman. Pompey took Jerusalem for Rome in 63 BCE; by the line’s last stretch the land was a client kingdom under Herod the Great and then a Roman province. It is into this people, in this province, at the turn of the era, that the body the next paper follows was born — a Jew of the tribe of Judah and, the tradition holds, of the house of David, the genealogies of Matthew 1 and Luke 3 tracing the descent down to him.
The fuller of the two, Matthew’s, is worth setting down whole, because the tradition did not merely assert the descent but counted it — and counted it in a deliberately patterned way, as three movements of fourteen generations each. The first runs from the patriarch to the king: Abraham, Isaac, Jacob, Judah, Perez (born of Tamar), Hezron, Ram, Amminadab, Nahshon, Salmon, Boaz (born of Rahab), Obed (born of Ruth), Jesse, and David the king. The second runs from the throne to the catastrophe: Solomon (born of the wife of Uriah), Rehoboam, Abijah, Asa, Jehoshaphat, Joram, Uzziah, Jotham, Ahaz, Hezekiah, Manasseh, Amon, Josiah, and Jeconiah, at the deportation to Babylon. The third runs from the exile to the child: Shealtiel, Zerubbabel, Abiud, Eliakim, Azor, Zadok, Achim, Eliud, Eleazar, Matthan, Jacob, Joseph the husband of Mary, and Jesus who is called Christ. Three fourteens, the line sounded as a measured descent, narrowing name by name to the one at its foot.
The patterning is exactly the place to be honest, and three things should be said plainly. Some of these names stand in the wider record beyond Matthew: the kings of the second movement are chronicled in Kings and Chronicles, and Shealtiel and Zerubbabel reappear in the books of the return (Ezra, Haggai) as real figures of the rebuilding. Some do not stand anywhere else: the stretch from Abiud to Matthan — the generations between the return and Joseph — is attested in no other source, biblical or external; these are Matthew’s names alone, a private line for a period the records otherwise leave dark. And some were, on the best reading, left out to make the count come right: to reach a clean three-times-fourteen, Matthew passes silently over three generations of Judah’s kings — Ahaziah, Joash, and Amaziah — between Joram and Uzziah, who stand in the Chronicler and are absent here. Matthew states the threefold-fourteen pattern himself (1:17); why fourteen, he does not say, and the honest reading leaves that open. A frequently cited suggestion ties the number to the name David, whose Hebrew letters (D-W-D, 4 + 6 + 4) sum to fourteen — but that is a proposal the text nowhere confirms, and the only thing the structure itself establishes is what the omissions already show: the line was arranged to carry a meaning, and a complete registry was not its first concern. The seam shows even in the counting, for the third movement as Matthew lists it holds only thirteen new names, the symmetry pressed a little harder than the bare arithmetic will bear. And Luke, tracing the same descent, runs it through a different son of David and gives Joseph a different father — the two genealogies agreeing at the ends and parting in the middle, as ancient genealogies kept for what they meant rather than for what a registrar would have certified often do. None of this unmakes the line; it only marks where the record is firm, where it is the tradition’s alone, and where the meaning was allowed to shape the count.
And here the long arithmetic of the paper closes. Two thousand years of one line, bodies begetting bodies from Abraham — and behind them the whole climb these pages have traced, the first particles that took on mass, the elements forged in dying stars, the first living cell, the four billion years of forms — all of it has been bearing toward one place, and here it arrives: a single body, in a single province, at the turn of the era. The one the whole climb was always going toward is here.
20. Born

The material thread does not end at a summit, and it does not, here, end at a grave. It ends at a beginning: a body, born — the same kind of body the whole climb has been assembling, hydrogen to stardust to the first cell to the primate line to the people of Israel, now drawn to a single point in a single infant, in a single province, at the turn of the era. By every measure this paper has used, it is an ordinary human body: the same chemistry, the same descent, the same four billion years behind it as any other body that has ever drawn breath. That is the whole weight of the claim, and the lens that follows the matter makes no larger one. What that body went on to do, and suffer, and become — the life it lived and the death it died — is not the material thread’s to tell; it belongs to the life, which is Paper A6. The long climb arrives at the cradle, sets down what it carried, and hands it on.
References
Allwood, A.C., et al. (2018). Reassessing evidence of life in 3,700-million-year-old rocks of Greenland. Nature 563, 241–244.
Araújo, R., et al. (2022). Inner ear biomechanics reveals a Late Triassic origin for mammalian endothermy. Nature 607, 726–731.
Bell, E.A., et al. (2015). Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon. PNAS 112, 14518–14521.
Black, B.A., et al. (2024). The anatomy and lethality of the Siberian Traps large igneous province. Annual Review of Earth and Planetary Sciences 52.
Bobrovskiy, I., et al. (2018). Ancient steroids establish the Ediacaran fossil Dickinsonia as one of the earliest animals. Science 361, 1246–1249.
Borsányi, S., et al. (2016). Calculation of the axion mass based on high-temperature lattice QCD. Nature 539, 69–71.
Brocks, J.J., et al. (2023). Lost world of complex life and the late rise of the eukaryotic crown. Nature 618, 767–773.
Bromm, V. (2013). Formation of the first stars. Reports on Progress in Physics 76, 112901.
Burbidge, E.M., Burbidge, G.R., Fowler, W.A., Hoyle, F. (1957). Synthesis of the elements in stars. Reviews of Modern Physics 29, 547–650.
Canup, R.M., Asphaug, E. (2001). Origin of the Moon in a giant impact near the end of the Earth’s formation. Nature 412, 708–712.
Carniani, S., et al. (2024). A shining cosmic dawn: spectroscopic confirmation of two luminous galaxies at z ≈ 14. Nature 633, 318–322.
Cazenave, M., et al. (2024). Postcranial evidence does not support habitual bipedalism in Sahelanthropus tchadensis: a reply to Daver et al. (2022). Journal of Human Evolution 192, 103557.
Chiarenza, A.A., et al. (2020). Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. PNAS 117, 17084–17093.
Connelly, J.N., et al. (2012). The absolute chronology and thermal processing of solids in the solar protoplanetary disk. Science 338, 651–655.
Cowan, J.J., et al. (2021). Origin of the heaviest elements: the rapid neutron-capture process. Reviews of Modern Physics 93, 015002.
Daver, G., et al. (2022). Postcranial evidence of late Miocene hominin bipedalism in Chad. Nature 609, 94–100.
During, M.A.D., et al. (2022). The Mesozoic terminated in boreal spring. Nature 603, 91–94.
Eme, L., Spang, A., Ettema, T.J.G., et al. (2017). Archaea and the origin of eukaryotes. Nature Reviews Microbiology 15, 711–723.
Fields, B.D. (2011). The primordial lithium problem. Annual Review of Nuclear and Particle Science 61, 47–68.
Fields, B.D., Olive, K.A., Yeh, T.-H., Young, C. (2020). Big-Bang nucleosynthesis after Planck. JCAP 03, 010.
Fischer, W.W., Hemp, J., Johnson, J.E. (2016). Evolution of oxygenic photosynthesis. Annual Review of Earth and Planetary Sciences 44, 647–683.
Gunz, P., et al. (2020). Australopithecus afarensis endocasts suggest ape-like brain organization and prolonged brain growth. Science Advances 6, eaaz4729.
Harmand, S., et al. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature 521, 310–315.
Hu, W., Dodelson, S. (2002). Cosmic microwave background anisotropies. Annual Review of Astronomy and Astrophysics 40, 171–216.
Imachi, H., et al. (2020). Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577, 519–525.
Javaux, E.J. (2019). Challenges in evidencing the earliest traces of life. Nature 572, 451–460.
Kasen, D., et al. (2017). Origin of the heavy elements in binary neutron-star mergers from a gravitational-wave event. Nature 551, 80–84.
Kobayashi, C., Karakas, A.I., Lugaro, M. (2020). The origin of elements from carbon to uranium. The Astrophysical Journal 900, 179.
Lane, N., Martin, W.F. (2012). The origin of membrane bioenergetics. Cell 151, 1406–1416.
Langergraber, K.E., et al. (2012). Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. PNAS 109, 15716–15721.
Lyons, T.W., Reinhard, C.T., Planavsky, N.J. (2014). The rise of oxygen in Earth’s early ocean and atmosphere. Nature 506, 307–315.
Mazumdar, A., White, G. (2019). Review of cosmic phase transitions: their significance and experimental signatures. Reports on Progress in Physics 82, 076901.
Moody, E.R.R., et al. (2024). The nature of the last universal common ancestor and its impact on the early Earth system. Nature Ecology & Evolution 8, 1654–1666.
Moorjani, P., et al. (2016). Variation in the molecular clock of primates. PNAS 113, 10607–10612.
Newham, E., et al. (2020). Reptile-like physiology in Early Jurassic stem-mammals. Nature Communications 11, 5121.
Nutman, A.P., et al. (2016). Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures. Nature 537, 535–538.
Patel, B.H., Percivalle, C., Ritson, D.J., Duffy, C.D., Sutherland, J.D. (2015). Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism. Nature Chemistry 7, 301–307.
Planck Collaboration (Aghanim, N., et al.) (2020). Planck 2018 results. VI. Cosmological parameters. Astronomy & Astrophysics 641, A6.
Poulton, S.W., et al. (2021). A 200-million-year delay in permanent atmospheric oxygenation. Nature 592, 232–236.
Prang, T.C., et al. (2021). Ardipithecus hand provides evidence that humans and chimpanzees evolved from an ancestor with suspensory adaptations. Science Advances 7, eabf2474.
Preiner, M., et al. (2020). A hydrogen-dependent geochemical analogue of primordial carbon and energy metabolism. Nature Ecology & Evolution 4, 534–542.
Riess, A.G., et al. (2024). JWST observations reject unrecognized crowding of Cepheid photometry as an explanation for the Hubble tension. The Astrophysical Journal Letters 962, L17.
Robertson, B.E. (2022). Galaxy formation and reionization: key unknowns and expected breakthroughs by JWST. Annual Review of Astronomy and Astrophysics 60, 121–158.
Rodrigues-Oliveira, T., et al. (2023). Actin cytoskeleton and complex cell architecture in an Asgard archaeon. Nature 613, 332–339.
Senel, C.B., et al. (2023). Chicxulub impact winter sustained by fine silicate dust. Nature Geoscience 16, 1033–1040.
Sevim-Erol, A., Begun, D.R., et al. (2023). A new ape from Türkiye and the radiation of late Miocene hominines. Communications Biology 6, 842.
Stewart, T.A., et al. (2020). The fin-to-limb transition reconstructed from the Tiktaalik pectoral fin. PNAS 117, 1612–1620.
Stockey, R.G., Sperling, E.A., et al. (2024). Life on the edge: the Cambrian marine realm and oxygenation. Annual Review of Earth and Planetary Sciences 52.
Trodden, M. (1999). Electroweak baryogenesis. Reviews of Modern Physics 71, 1463–1500.
Valley, J.W., et al. (2014). Hadean age for a post-magma-ocean zircon confirmed by atom-probe tomography. Nature Geoscience 7, 219–223.
Villmoare, B., Delezene, L.K., Rector, A.L., et al. (2025). New discoveries of Australopithecus and Homo from Ledi-Geraru, Ethiopia. Nature.
Vosseberg, J., et al. (2025). Dated gene duplications elucidate the evolutionary assembly of eukaryotes. Nature.
White, T.D., et al. (2009). Ardipithecus ramidus and the paleobiology of early hominids. Science 326, 64–86.
Wilson Mantilla, G.P., et al. (2021). Earliest Palaeocene purgatoriids and the initial radiation of stem primates. Royal Society Open Science 8, 210050.
Zhu, M., et al. (2022). The oldest complete jawed vertebrates from the early Silurian of China. Nature 609, 954–958.
Added for §17 (the behavioral record of consciousness):
Henshilwood, C.S., et al. (2002). Emergence of modern human behaviour: Middle Stone Age engravings from South Africa. Science 295, 1278–1280.
McBrearty, S., Brooks, A.S. (2000). The revolution that wasn’t: a new interpretation of the origin of modern human behavior. Journal of Human Evolution 39, 453–563.
Added for §§18–19 (the peopling and the line of Israel):
Dever, W.G. (2003). Who Were the Early Israelites and Where Did They Come From? Eerdmans. [A centrist synthesis of the archaeology of early Israel.]
Finkelstein, I., Silberman, N.A. (2001). The Bible Unearthed: Archaeology’s New Vision of Ancient Israel and the Origin of Its Sacred Texts. Free Press. [A minimalist-leaning synthesis; cited for the live debates, not as the settled view.]
Added for §§20–21 (the human body and the physiology of the death):
Barbet, P. (1953). A Doctor at Calvary: The Passion of Our Lord Jesus Christ as Described by a Surgeon. P.J. Kenedy. [The asphyxia hypothesis; a primary source for one side of the debate, not a current consensus.]
Edwards, W.D., Gabel, W.J., Hosmer, F.E. (1986). On the physical death of Jesus Christ. JAMA 255, 1455–1463.
Haas, N. (1970). Anthropological observations on the skeletal remains from Giv’at ha-Mivtar. Israel Exploration Journal 20, 38–59.
Hublin, J.-J., et al. (2017). New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature 546, 289–292.
Maslen, M.W., Mitchell, P.D. (2006). Medical theories on the cause of death in crucifixion. Journal of the Royal Society of Medicine 99, 185–188.
Zias, J., Sekeles, E. (1985). The crucified man from Giv’at ha-Mivtar: a reappraisal. Israel Exploration Journal 35, 22–27.
Zugibe, F.T. (2005). The Crucifixion of Jesus: A Forensic Inquiry. M. Evans.